Immune responses to viruses
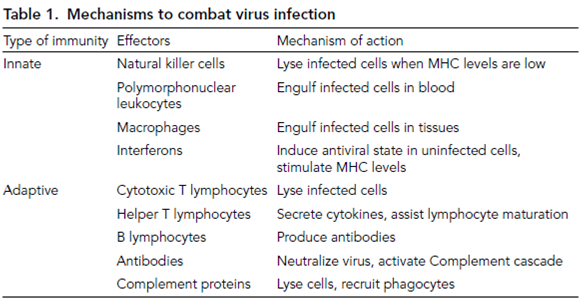
The immune system has two types of effectors; those of the innate response, which act early during infection in a general, nonspecific manner, and the cells of the adaptive response, which are activated a few days later and react specifically with the stimulat- ing antigen. The innate immune system responds first in combating virus infections and comprises lytic (natural killer, NK) cells and phagocytic cells that are found either within the blood (polymorphonuclear leukocytes, PMNLs) or that can trawl the tissues (mac- rophages). NK cells have an important role in the control of some virus infections. In addition, infected cells themselves can respond to virus by the synthesis of interferon, a secreted protein that can induce an antiviral state in other, as yet uninfected cells, protecting them from viral invasion. Interferon proteins can also enhance the activity of immune lymphocytes.
The adaptive response requires several days to activate and consists of the T lymphocytes and, a few days later, the B lymphocytes. Before activation, the numbers of these cells are low but recognition of viral antigens stimulates expansion of activated cell lines that give rise to either undifferentiated memory cells or differentiated effectors. T lym- phocytes recognize viral antigens (via the T-cell receptor, TCR) that have been processed and exposed on the surface of cells in conjunction with host major histocompatibility complex (MHC) proteins. There are two types of T lymphocytes important in viral infections and they recognize antigens in association with different MHC proteins. Cytotoxic T lymphocytes (CTLs, also known as T8 or CD8 cells) lyse infected cells following the binding of viral antigens in association with MHC type I proteins that are found on all cells except neurons and red blood cells. Helper T lymphocytes (T4 or CD4 cells) bind viral antigens in association with MHC type II proteins that are found only on certain antigen-presenting cells (APCs) including dendrites and macrophages. These lympho- cytes are crucial for the action of both CTLs and B lymphocytes through the production of cytokines, soluble proteins that enhance or suppress immune cell functions. B cells similarly recognize viral antigens via a cell surface receptor (the B-cell receptor, BCR) and differentiated lymphocytes (plasma cells) synthesize antibody that binds with the same specificity (i.e. to the stimulating antigen). Antibodies may bind with antigen present on free virions or on infected cell surfaces. The binding of antibody to free virus can neutralize its infectivity by blocking virus attachment, penetration or uncoating. Binding of antibody to infected cells triggers the action of the Complement cascade, a series of nine soluble proteins that assemble into a pore structure (the membrane attack complex) that leads to infected cell lysis. The presence of antigen-bound antibody or Complement molecules on the surface of an infected cell can also recruit phagocytic cells (a process known as opsonization) to engulf the cell.
For acute infections, initial (primary) T-cell responses usually peak between 7 and 10 days post infection and decline within 3 weeks. The primary antibody response usually peaks later and is often barely detectable during the acute stage of infection but serum antibody levels increase dramatically 2–3 weeks post infection and may linger for several months. Primary antigen recognition and immune responses usually result in the formation of memory B and T cells that persist, making antigenic recognition much faster during reinfection. This secondary response is virtually immediate in terms of CTL and antibody response and often at a higher level. The mechanisms that combat viral infec- tions are outlined in Table 1.