The lac operon
The most generally used model for transcription and the regulation of transcription is the lac operon of E. coli. The operon comprises a promoter an operator a site on the DNA where regulatory proteins bind and three genes. These three genes allow the Bacterium to use lactose instead of glucose as a carbon source in Figure lacZ coding for the enzyme β-galactosidase the gene product LacZ – note the difference in italicization and capitalization between the gene and the protein it makes lacY coding for a permease which allows lactose through the membrane and lacA a gene of poorly understood function which codes for a transacetylase.
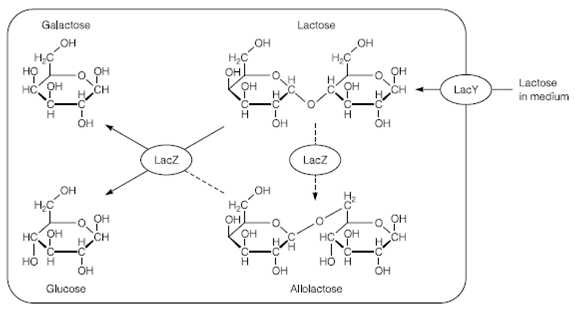
Figure: Lactose metabolism in Escherichia coli.
Upstream of the lacZYA promoter is another gene called lacI which has its own promoter in Figure. The product of this gene the protein LacI is the primary regulator of the lac operon and is sometimes called the Lac repressor. When LacI is bound to the lacZYA operon operator transcription is blocked the cell is unable to produce b-galactosidase and therefore is unable to use lactose as a carbon source.
The functions of all the genes were found as a result of analysis of mutations. Changes in the DNA sequence in the genes themselves can knock out individual genes in Table 1. The finding of a mutation which meant an inactive LacI was produced and this allowed constitutive expression of lacZ example for in the presence and absence of lactose was significant and was one finding that allowed Jacob and Monod to start to elucidate the mechanism of LacI repression. They proposed that LacI was always produced from its own promoter but was structurally altered through the presence of lactose itself that stopped it binding to the lacZYA promoter in Figure. It was found that the molecule that caused this change and induced the lac operon was allolactose in Figure. Allolactose is the primary inducer but there are other molecules like as XGAL which can also induce the operon.
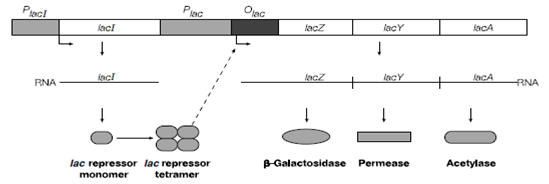
Figure : Structure of the lactose operon
Table 1. Effect of mutations in lac operon structural genes
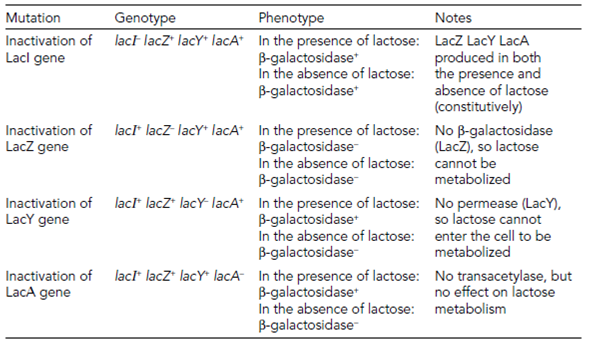
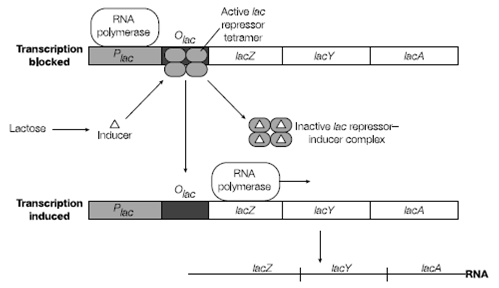
Figure: Binding of the inducer activates the lac repressor.
The model was backed up with studies in the nonstructural parts of the operon in the promoters of both lacZYA and lacI and the operator of lacZYA in Table 2.
The lac promoter itself is relatively weak even when induced by allolactose. High levels of transcription are only achieved when not only is the promoter derepressed but also when it is activated through the cAMP receptor protein. This is sometimes called catabolite activator protein CAP. If the cell is rich in energy from other sources, it may not need to utilize lactose at all. The precursor of ATP and AMP is in equilibrium with a cyclic form cAMP in there is a lot of cAMP. The cAMP binds to CRP which can then activate the lac operon through as much as 40-fold.
Figure: When glucose is low there is an abundance of AMP which in turn means
Table 2. Effect of mutations in lac operon promoters and operators
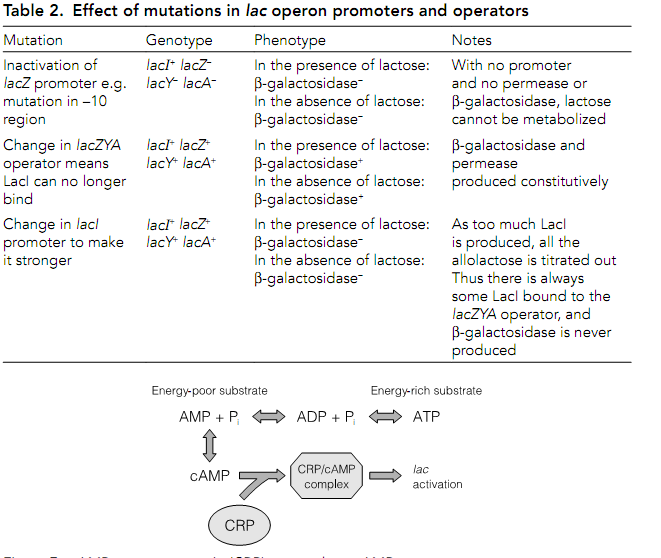
Figure: cAMP receptor protein (CRP) responds to cAMP.