Parallel processing in v1
The three relatively independent pathways, each of which processes the various aspects of vision in parallel can be delineated in the main visual cortex.
The magnocellular pathway from M ganglion cells to the M LGN cells has its input to spiny stellate cells in layer 4Cα. Such excitatory interneurons synapse with pyramidal cells in layer 4B that show orientation and direction selectivity. These cells send axon collaterals to the pyramidal cells in layers 5 and 6. The layer 5 cells project to subcortical areas, the pulvinar (a thalamic nucleus included in visual attention), the superior colliculus, and pons. The layer 6 pyramidal cells go to the extrastriate cortex. M pathway is an expert for analysis of motion. Its outputs through layer 5 are significant in visual attention and gaze reflexes. A few cells in the M pathway are binocular therefore it contributes to stereopsis. Since it originates with ganglion cells that combine input from the two classes of cone cell it is not wavelength selective; that is the M system is color blind.
There are two parvocellular pathways. They begin from P ganglion cells through P LGN cells that synapse with spiny stellate cells in 4Cβ. Similar to the M pathway the interneurons connect with pyramidal cells in 4B. Though, in the parvocellular paths, 4B cells (that are orientation-selective simple cells) synapse with the pyramidal cells in layers 2 and 3 which then relay with the deep pyramidal cells in layer 5. The Segregation of the two parvocellular pathways takes place in layers 2 and 3. Whenever stained for the mitochondrial enzyme, the cytochrome oxidase, layers 2 and 3 display pillars of high activity, blobs. Each of the blobs is centered on an ocular dominance column. Between the blobs lies the interblob area. Cells in the interblob area are orientation selective, binocularly driven, the complex cells. They are not motion sensitive or wavelength selective. They are section of the parvocellular–interblob (PI) pathway that processes high resolution analysis of form in the visual world. By contrast the cells in blobs are wavelength selective, display poor orientation selectivity and are monocular. The parvocellular–blob (PB) pathways mediate color vision. The blob pyramidal cells get direct input from blue-on K cells in the koniocellular LGN layers; however the function of this input is not yet understood.
The wavelength selective blob cells are double opponent cells with the receptive field (RF) properties derived from their inputs that is the single opponent parvocellular LGN cells. The double opponent cells have center–surround antagonist RF configuration, they signal the color contrast and come in four classes classified by their preferred stimuli. The left cell in figure shown below is excited by L cones in the center and inhibited by L cones in the surround. Additionally, it is inhibited by M cones centrally but excited by M cones in the surround. The favored stimulus for this cell is a red spot in a green background. Though, the cell gives off-responses when exposed to a green spot in a red background as shown in next figure.
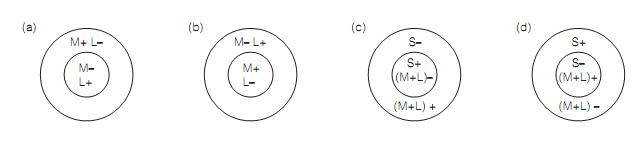
Figure: Double opponent cells in V1 blobs. Preferred stimuli: (a) red spot-green surround; (b) green spot-red surround; (c) blue spot-yellow surround; (d) yellow spot-blue surround.

Figure: Responses of the double opponent cell in previous figure: (a) preferred stimulus; (b) off response that may account for successive color contrast.
Dissimilar single opponent cells that are excited by small spots of white light, double opponent cells are unaltered by the white light stimuli of any size; therefore they are more selective detectors of the color contrast. The association of double opponent cell RFs elucidates some of the properties of color vision.
- Color constancy: The way in which the brain evaluates color constancy is not understood in detail but is partially accounted by the behavior of the double opponent cells. A shift in wavelength composition of light will produce equivalent but opposite effects on the responses of the center and surround of double opponent cells. There will be a minute effect overall on the RF of the cell that will continue to signal similar color. On the scale of the whole visual field, the color constancy is thought to include comparing red–green brightness, blue–yellow brightness, and total brightness (added outcomes of S, M, and L cones) over large regions of retina.
- Perceptual cancellation is elaborated by the manner in which the color opponency occurs to be organized as red (R) versus green (G) and yellow (R+G) versus blue channels. As mutual antagonism takes place among red and green or among yellow and blue only one color in each of the pair can be seen at a single pixel of retina at any time.
- Simultaneous color contrast can also be accounted for by the properties of double opponent cells. For illustration, the cell in figure above cannot differentiate between a green stimulus to its surround and a red stimulus to the center; the response is similar for both. Therefore a gray disc viewed in a green background is interpreted as red. The similar mechanism elucidates the complementary after-images which appear after staring at a uniform patch of color as shown in the figure above.