Carbon and energy metabolism
All species of the Archaeplastida, and many species of excavates and chromalveolates are photosynthetic and therefore gain carbon and energy from the fixation of atmospheric or dissolved carbon dioxide using photosynthesis.
The photosynthetic reactions take place in the chloroplasts, the light reactions occurring in the chloroplast thylakoids, and the light-independent reactions occur- ring in the stroma. The chlorophyll pigments are membrane-bound within the thylakoids, and their properties include the ability to be excited by light. Different chlorophylls accept light energy of different wavelengths, depending on their structure. Accessory pigments, the carotenoids, phycobilins, and xanthophylls, also absorb light energy of different wavelengths and pass their excitation to chlorophyll, maximizing the breadth of wavelength over which light energy can be absorbed. The role of accessory pigments is particularly important as they allow photosynthetic organisms to occupy different parts of the photic zone, the shallow layer of water where sufficient light penetrates to support photosynthesis. The actual depth to which light will penetrate varies with turbidity and dissolved organic matter content. Furthermore, light of different wavelengths penetrates water to different degrees. Red light is absorbed rapidly by water, blue light least. The members of the green algae occupy only the shallowest of waters.
Chlorophylls a, b, and c are the photosynthetic pigments, with chlorophyll a being the primary photosynthetic pigment. The absorption maxima of chlorophyll a are at 430 and 663 nm. Other chlorophylls are accessory pigments and have slightly different absorption maxima: chlorophyll b at 435 and 645 nm, chlorophyll c1 at 440, 583, and 634 nm, and c2 at 452, 586, and 635 nm. Phycobilins absorb light energy at 565 nm. There are also other accessory pigments found in the different algal groups, the carotenes and xanthophylls. Depending on the different accessory pigments present in individual species, light of different wavelengths can be absorbed and utilized in photosynthesis.
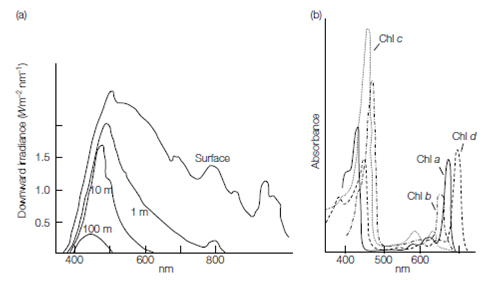
Figure 1. (a) Downward penetration of different light wavelengths. (b) Absorption spectra of chlorophylls.
Nutrient uptake in nonphotosynthetic excavates and chromalveolates may be by diffusion, pinocytosis, or phagocytosis. Within nutrient-rich tissues, such as those that intra- cellular parasites or blood-dwelling parasites live in, nutrient uptake is by diffusion and pinocytosis, the formation and digestion of small vesicles from the plasma membrane. These vesicles capture soluble nutrients from the environment.
In larger, free-living species, particulate organic matter and bacteria are engulfed by phagocytosis. In this process large vacuoles can form anywhere over the cell surface in the amebae, or from specialized sites such as the cytostome in ciliates and flagellates. Digestion of the vesicle content occurs by the fusion of lysozyme-containing enzyme vesicles with the phagocytic vesicles. Once digestion is complete enzymes are recycled by the cell and sequestered into small vesicles, while cell debris is expelled from the cell by reverse phagocytosis, where the food vacuole re-fuses with the plasma membrane.
A large number of symbiotic species can be found in the excavates and chromalveolates in association with many different photosynthetic symbionts. Species of Alveolates and Euglenoids with these symbionts have an autotrophic nutrition, relying on photosynthetic products for carbon and energy sources. In some cases the cytostome seen in nonphotosynthetic protistans is not present. Further evidence of the close taxonomic relationships between the photosynthetic and nonphotosynthetic groups is provided by the fact that it is possible to ‘cure’ some species of their symbionts, and once cured, these protista return to their holozoic nutrition and form food vesicles from a reformed cytostome.
Almost all members of the green algae are aerobic and use mitochondrial respiration with oxygen as the terminal electron acceptor. In most of the archaeplastids, excavates, and chromalveolates, glycolysis and the citric acid cycle provide energy and intermediates for cellular metabolism. Aerobic species use mitochondrial respiration, with oxygen as the terminal electron acceptor, generating ATP.Anaerobic species have a fermentative metabolism, which results in the incomplete oxidation of substrates. A few species that live in anaerobic lake sediments can use alternative electron acceptors such as nitrate.
There are several modifications to the usual glycolytic pathway found in some members of excavates and chromalveolates. Some can utilize inorganic pyrophosphate rather than ATP replacing enzymes like pyruvate kinase with pyrophosphate kinase. The advantage of this is that pyrophosphate kinase activity is reversible and can be used to synthesize glucose from other substrates.In parasitic kinetoplastids glycolysis does not occur in the cytoplasm but in its own organelle, the glycosome . Within this organelle a modified form of glycolysis can give rise to glycerol, which is the substrate of respiration in this group. In anaerobic species fermentative reactions occur within the hydrogenosome. Substrates like pyruvate and malate from glycolysis are taken up by this organelle and incompletely oxidized to end products like acetate, in the process forming ATP. Hydrogen is another characteristic end product of hydrogenosomal metabolism.
In many excavates and chromalveolates, mitochondrial metabolism resembles that of bacterial respiration, having a branched pathway, part of which is sensitive to cyanide (mammalian-like) and part of which has an alternative terminal oxidase that allows the mitochondrion to operate at low oxygen levels. These modifications are most developed in the specialized parasites, where both cellular and mitochondrial structure change as the parasite passes through different stages of the life cycle.
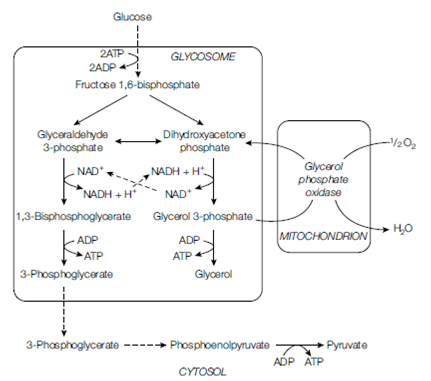
Figure: Aerobic glycolysis in bloodstream forms of trypanosomes, showing compartmentalization of much of the pathway in the glycosome. Dotted lines indicate several reaction steps.