Primary visual cortex
The fibers of the optic tract finished in the striate cortex (Brodmann’s region 17) on the medial surface of the tip of the occipital lobe. This area is the primary visual cortex (V1). Accurate retinotopic mapping is maintained up to V1 with the fovea having a disproportionately big representation.
There are at least three parallel streams of information into the primary visual cortex. Movement sensitive M LGN cells input into layer 4Cα, the P LGN cells go to layer 4Cβ, while the koniocellular layers of the LGN project to layers 2 and 3. Such streams remain quasi-independent throughout the visual system. The connections of primary visual cortex are illustrated for the primate in as shown in figure below:
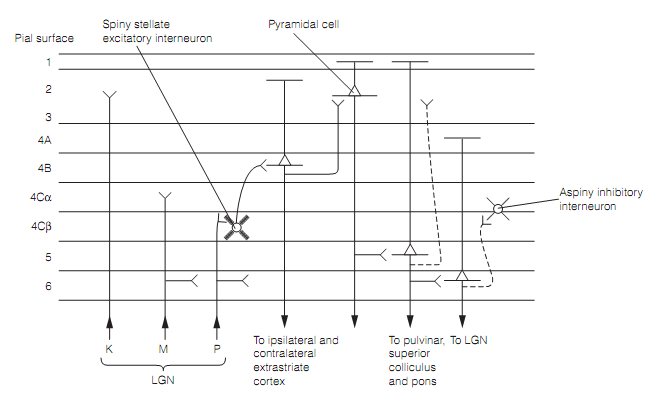
Figure: Canonical circuitry in the primary visual cortex illustrated for parvocellular (P) LGN input. Magnocellular (M) circuitry has its input to 4Cα.
The great majority of cells in V1 has extended receptive fields (RFs) with both inhibitory and excitatory regions, and responds to slits, bars, edges, and corners instead spots of light. Most fall into two class based on their RF properties: simple or complex cells. Both are orientation selective, so they react to linear features in only a narrow variety of orientations:
- Simple cells are pyramidal cells found generally in layers 4 and 6. They are highly sensitive to the location of a stimulus on the retina. They contain small oval RFs with center–surround antagonism as shown in figure. A simple cell gets its input from a linear array of LGN cells having similar RF properties, therefore the RF of the simple cell emerges as a result of the RFs of the LGN inputs.
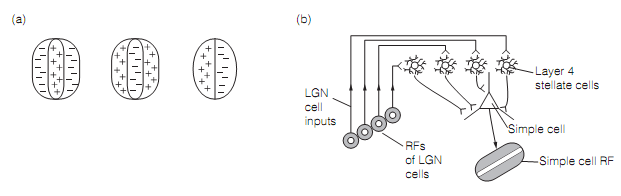
Figure: (a) Receptive fields of three simple cells; (b) a diagram depicting how lateral geniculate nucleus (LGN) cells contribute to the simple cell receptive field (RF), four on-center LGN cell RFs generate an on-center simple cell RF.
- Complex cells are most abundant in layers 2, 3, & 5. They have larger RFs as compare to simple cells and, lacking different inhibitory or excitatory areas, a stimulus of the suitable orientation anywhere in the RF evokes a response. Therefore, complex cells are much less fussy about location than simple cells. Most complex cells show a preference for movement at right angles to the long axis of the stimulus. A few complex cells receive their inputs from simple cells however others get their input directly from the LGN.