Motor cortex
The motor cortex is subcategorized into three areas, MI, MII that contains the supplementary motor area (SMA), and premotor area (PM). These are reciprocally connected with each other as shown in figure and with sub-cortical structures that send inputs back to the motor cortex through the thalamus forming closed motor loops. Reciprocal reverse projections from the motor cortex to the thalamus also exist.
The supplementary motor area is portion of a motor loop with the basal ganglia. It sends output to the striatum that projects back to the SMA through the globus pallidus and ventrolateral (VLO) thalamus. Most corticospinal tract axons from M1 either terminate in the pons or give off collaterals there. These build synapses with pontine neurons that project to the cerebellum. Outcomes from the cerebellum to the thalamus that in turn project back to M1 and PM motor regions form other motor loops. These motor loops by basal ganglia and cerebellum are needed for the initiation of particular motor patterns and their coordination.
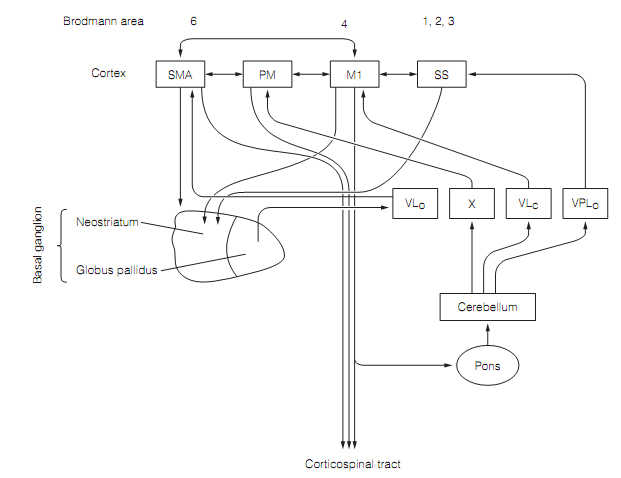
Figure: The connections of the motor cortex begin motor loops with the basal ganglia and the cerebellum in which the thalamus gives the input which closes the loop. Here SS=somatosensory cortex. Thalamic nuclei: VLc= ventroposterior nucleus pars caudalis; VLo=ventroposterior nucleus pars oralis; VPLo=ventroposterolateral nucleus pars oralis; X= nucleus X.
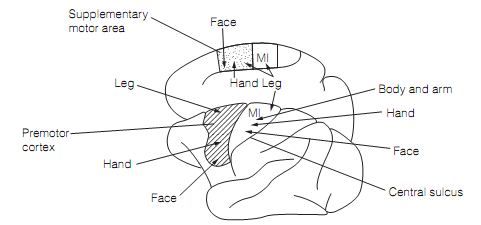
Figure: Approximate somatotopic mapping in the motor cortex of the macaque. Here M1=primary motor cortex.
Numerous somatotopic maps exist in the motor cortex as shown in figure. Their topography is preserved in the orderly arrangement of the fibers in the corticospinal tract. Similar to somato-sensory maps, such motor maps are grossly distorted. More cortical space is dedicated to the tongue, face, and hands than other areas, permitting great range and precision of movements. The M1 obtains a substantial input from the somatosensory cortex and many M1 neurons have sensory receptive fields. These are situated where activity of the neuron is likely to cause movement; that is, the M1 neurons are wired to be responsive to the sensory consequences of their proceedings.
What the somatotopic mapping in the motor cortex stands for is not certain. It is not a one-to-one mapping to separate muscles or movements. This is shown as follows:
? The output from single cortical neurons diverging to numerous motor neuron pools
? Converging outputs from quite a broad region of M1 onto the motor neuron pools for muscles moving a particular body part.
? A given muscle being subserved by an area in the motor cortex that overlaps with areas controlling neighboring muscles.
During the execution of a movement including a muscle the set of neurons in the area which are activated depends on the nature of the movement; for illustration, its direction and force.
Recording from single cells in the motor cortex of conscious monkeys engaged in intentional limb movements illustrates that M1 cell firing can correlate with velocity, force, rate of change of force, direction of movement, acceleration, or joint position. None of these parameters is mapped in an orderly manner in the cortex. Firing of an M1 cell during a task is generally associated to two or three of these variables therefore M1 cells do not completely encode a single movement parameter. Most M1 cells are instead broadly tuned for movement direction therefore a single M1 cell cannot encode direction of movement very well. Though, direction of movement is very accurately coded by the average firing of a few hundred cells. This is an illustration of population coding. The population of cells encoding the way of a given movement is quite broadly distributed across the cortex.
The secondary motor cortex consists of neurons which fire in a manner which correlates with the direction and force of a movement. The SMA control proximal limb muscles directly through its outcome to the corticospinal tract, while the premotor region neurons synapse with brainstem reticular neurons which go to axial and proximal limb muscles. Both control distal limb muscles through M1.
The SMA has bilateral presentations of the body and is vital for movements including both sides of the body, mainly those that have been learnt. For this, the motor loop including the basal ganglia is significant. The premotor region consists of a number of discrete motor areas and therefore has numerous maps. It is implicated in movements in response to sensory cues. The premotor region gets a large input from the posterior parietal cortex (Brodmann regions 5 and 7) relationship cortex that receives visual, somatosensory, and the vestibular sensory input. The posterior parietal cortex therefore gives sensory input for targeted movements. Few posterior parietal neurons are context precise, firing only during goal-directed behavior (example, reaching for food) although remaining silent if the limb moves in similar way in the absence of the aim.
A key role of the secondary motor cortex is in planning movements. This is shown, at first, by the fact that neurons here fire a long time (probably up to 800 ms) before a voluntary movement starts. Secondly, measurements of cerebral blood flow (cbf ) in humans doing motor tasks illustrate that a simple movement includes increased cbf in MI only, a more difficult task is accompanied by increased cbf in the secondary motor cortex and also MI; although, remarkably, whenever subjects were needed to mentally rehearse a complex task (though not do it) an increase in cbf was viewed limited to the secondary motor cortex.