Feedback mode
During the implementation of well-rehearsed movements which are not too rapid the cerebellum acts as a feedback device to compare motor intentions with motor performance, and works to decrease any mismatch among them. For the spinocerebellum, the motor intentions are the signals transmitted by the corticopontine-cerebellar tract. Motor performance is monitored by proprioceptor input, and by the ventral spinocerebellar signals reporting on the activity of spinal cord and brainstem motor circuits. Likewise, the cerebrocerebellum compares inputs from the supplementary motor cortex and the primary motor cortex to generate error signals which reflect a discrepancy among motor commands and motor planning. In each situation the error signals are used to correct the mismatch.
The intermediate spinocerebellum seems to be included in correcting mistakes in limb movements like whenever a limb is perturbed by unexpected force. The order in which different neural elements fire in this situation is: muscle afferents–interpositus nucleus–motor cortex–dentate nucleus.
Feedback error correction perhaps works as follows. An error means that the real location of a limb is not the intended one. This generates unpredicted muscle stretch as the suitable coactivation of α and γ efferents to the muscle spindle have not occurred. Accurately the similar thing occurs when an unexpected force is applied to a limb. In either situation the muscle stretch will excite Ia and Ib afferents in the loaded muscles. These proprioceptor signals are transmitted to the cerebellum by mossy fibers. The stimulated mossy fibers bring tonic excitation to the intracerebellar nucleus through axon collaterals and stimulate a group of granule cells. The granule cell parallel fibers trigger numerous arrays of on-beam Purkinje cells as shown in figure below which strongly inhibit their target neurons in the interpositus nucleus. Generally, whenever mossy fibers are firing at background rates their tonic facilitation of the interpositus dominates the inhibition by Purkinje cells. Accordingly, interpositus neurons sustain excitation of the ventrolateral thalamus and the red nucleus. Whenever the mossy fibers are activated during a movement, though, Purkinje cell inhibition dis-facilitates the interpositus neurons and this inhibition is transmitted downstream to the red nucleus and thalamus. In contrary, neighboring off-beam PC arrays, reserved by the GABAergic interneurons in the cortex permit their interpositus cells to fire at higher than background rates. Therefore, the pattern of activation of the deep cerebellar nucleus is a negative image of the input activation.
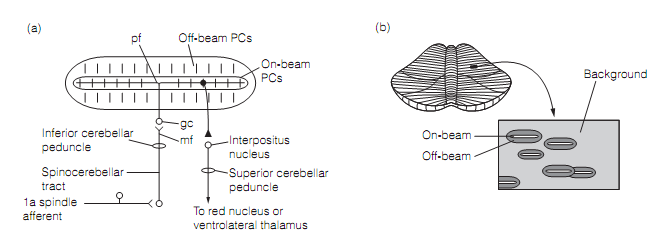
Figure: (a) Mossy fiber (mf) input activates an array on on-beam Purkinje cells (PCs). Each little vertical line represents the planar dendritic field of a PC, viewed from top. The output of only one PC is shown. (b) The Pattern of activation produced by single mossy fiber input. Here, gc=granule cell; pf=parallel fiber.
The total effect, mediated by the rubrospinal and corticospinal tracts, is to right the movement error by activating spinal reflexes which defend the correct limb position and dampen those that do not. This is probably completed by adjusting the firing of γ efferents to muscle spindles.
The intermediate spinocerebellum appears to control the precise timing of the contraction of agonist and antagonist muscles during a movement. During the reciprocal activation of agonist and antagonist muscles, Purkinje cells are responsible for controlling such muscles fire alternately, driving interpositus neurons to do the similar. During co-contraction, though, the Purkinje cells are silent. A role for the cerebellum in systematizing these patterns of muscle activity is supported by the fact that the action tremor resultant from lesions of the interpositus nucleus or intermediary cerebellar cortex seems to be due to derangement in the timing of agonist contraction. In average humans, a rapid wrist movement includes an initial burst of activity in the agonist, followed by burst in the antagonist to generate braking, and finally a second agonist burst to stabilize the joint at the preferred end point. In cerebellar tremor the beginning of the movement is normal but the second agonist burst is late. Accordingly the antagonist burst moves the wrist beyond the finish point. This sets up the tremor.